新しい視点で魚を科学する 水産生物学研究室
染色体操作研究CHROMOSOME Manipulation FOR fISH BREESSING
| 染色体操作とは |
| 遺伝子をゲノム単位で増やしたり、減らしたり、違うものを組み合わせたりして、有用な特性を引き出す育種法です。もともとその種がもっている遺伝子をそのまま使います。ゲノムとは生物の生存に必要な遺伝子群をもったひと組の染色体のセットのことで、通常の動物はこれを2セットもっているので2倍体といいます。これを3セット、4セットもっているのが3倍体、4倍体・・・・。これらを倍数体とよびます。こういう倍数体を作出することで生物の特性を改良し利用することを倍数性育種といいます。 |
初期の細胞分裂を操作する−倍数体、雌性・雄性発生二倍体−
染色体操作によって作出できるのは、三倍体triploidや半数体haploid、四倍体tetraploid、雌性発生二倍体gynogenetic diploidや雄性発生二倍体androgenetic diploidなどがあります。また異種間交雑を利用すれば、異質三倍体allotriploidや複二倍体amphidiploidをつくることができます。いずれも、受精前後の初期発生段階で、紫外線などをつかって染色体を働かなくしたり、高温や低温、圧力をつかって細胞分裂を阻止したりしてつくります。染色体操作法の技術開発はその強さやタイミングについて最適な条件を探す研究です。魚卵の染色体の倍数化は、主に第2減数分裂と卵割のいずれかを阻止することを狙って行いますが、同じ細胞分裂阻止でも、時期によって作出できるものが違ってきます。通常の受精卵の第2減数分裂を阻止すると三倍体が、第1卵割を阻止すると四倍体がそれぞれ作出されることになります。
純系育種のための雌性発生卵の卵割阻止の簡易化−高温刺激での中心小体の破壊を制御する−

単性発生卵(雌性発生卵や雄性発生卵)の卵割を阻止して作出される倍加半数体(doubled haploid, DH)は、すべての遺伝子座がホモ接合になるため、卵や精子をつくる配偶子形成で減数分裂の影響を受けず、つくられるすべての配偶子の遺伝子組成が同じになります。そのためにDHの次世代を再度単性発生で作出すると、そのきょうだいたちはすべての遺伝子が同じクローン集団になります。つまり、この方法を使うと、たった2世代で純系を作り出すことができます。これまで、この方法をつかって、様々な魚種で純系の作出が試みられてきました。 ところが、この卵割阻止はとても難しく、たくさんのDHを一度に生産することができません。とりあえず、クローン集団はつくれるようになったのですが、優良な特性をもったクローン系統を作出するには、たくさんのDHを生産し、様々な特性について個体ごとに比較して、優秀な成績を示すDHを選抜できるようにする必要があります。DHをもっと簡単に効率よくつくるにはどうすればよいか。それが今の最も大きな課題です。
卵割阻止が簡単になれば、多くのDHが得られるはずです。それには卵割のメカニズムを解明することが重要です。卵割阻止を行うときには卵を高水圧にさらしたり、温水や冷水につけたりといった物理的刺激処理をつかいます。これらの刺激処理を行うと、何がおきるのでしょうか。卵の細胞質の中には微小管という糸状の構造体があって、これが染色体を動かします。その微小管は中心体という細胞内小器官から伸びてきます。以前は、この微小管を破壊すれば染色体の分離がとまり、細胞分裂が阻止されると考えられていました。ところが、卵割のときにこの微小管を破壊しても、しばらくすると中心体からまた微小管が再生し、結局卵割は起こってしまうことが多いのです。ところが、水圧処理をすると卵割阻止がうまくいくこともありました。
この中心体の中には、中心小体というものが通常は1対ずつあり、それぞれ母中心小体と娘中心小体といいます。これらは親子関係で、娘中心小体は母中心小体から複製されてつくられます。第一卵割で高水圧などの物理的刺激を受けると、この中心小体が破壊されることがあり、特に娘中心小体が母中心小体より破壊されやすいそうです。2つの中心体の中でそれぞれの娘中心小体が破壊されると、母中心小体だけになるのですが、この状態でも中心体として微小管をつくることはできるので、微小管でつくられる紡錘体は再生され、第一卵割は正常に進行します。しかし、そこから次の卵割に入る前、卵は複製期にはいります。通常の場合、複製期では染色体のDNAが複製して2倍になったり、中心体も複製して2倍になったりします。通常の中心体では中心体の中の母娘中心小体が分かれて、それぞれが自分の娘を複製し、2個の完全な中心体になるのです。しかし、娘中心小体のない中心体では母中心小体だけが複製して自分の娘をつくるので、中心体としての数は増えません。中心体は紡錘体をつくるかなめとなるので、これが細胞あたり2個に増えないと、細胞分裂をおこなうための極は1個しかできず、複製されて2倍になった染色体は分離することなく1つの極にあつめられ、1つの大きな核を作ります。これが卵割阻止のメカニズムです。
中心小体の壊れ方は様々なパターンが考えられます。片方の中心体の娘中心小体だけが壊れる場合から、両方の中心小体がそれぞれ壊れ結果として片側の母中心小体しか残らない場合もあるかもしれません。それぞれで様々な壊れ方をすると、その後の分裂の仕方はさまざまなことが起こり、4極性や3極性の分裂をしたり、第一卵割が阻止されたり、第一卵割はそのまま起こって第2卵割が阻止されることもあるかもしれません。このような中心小体の破壊をうまく制御して、どの卵でも同じように卵割がうまく阻止されるような方法を開発することがとても重要です。
性成熟の弊害を防止してオールシーズンで肉質を維持する
ー三倍体と不稔性−

たとえばニジマスやサケ、アマゴ、ヤマメなどのサケ科魚類では、秋になると体中の栄養を卵巣や精巣に集中することで肉質が落ち、また婚姻色で体色が黒ずみ商品価値が大きく下がります。また、サケやアマゴのほとんどの親は産卵で体力を使い果たして死んでしまいます。
三倍体はゲノムを3組もつので、すべての相同染色体が3つずつあり、減数分裂がうまくいかないことからその後の生殖巣の発達が抑制されるので、産卵期に卵に栄養をとられ肉質が落ちる季節にも若々しい肉質を保つことができるし、産卵したら死んでしまうような種では性成熟の抑制によって通常よりも長く生きることができます。
雄親の遺伝的影響を除外して優良な系統をつくる ー雌性発生育種法−
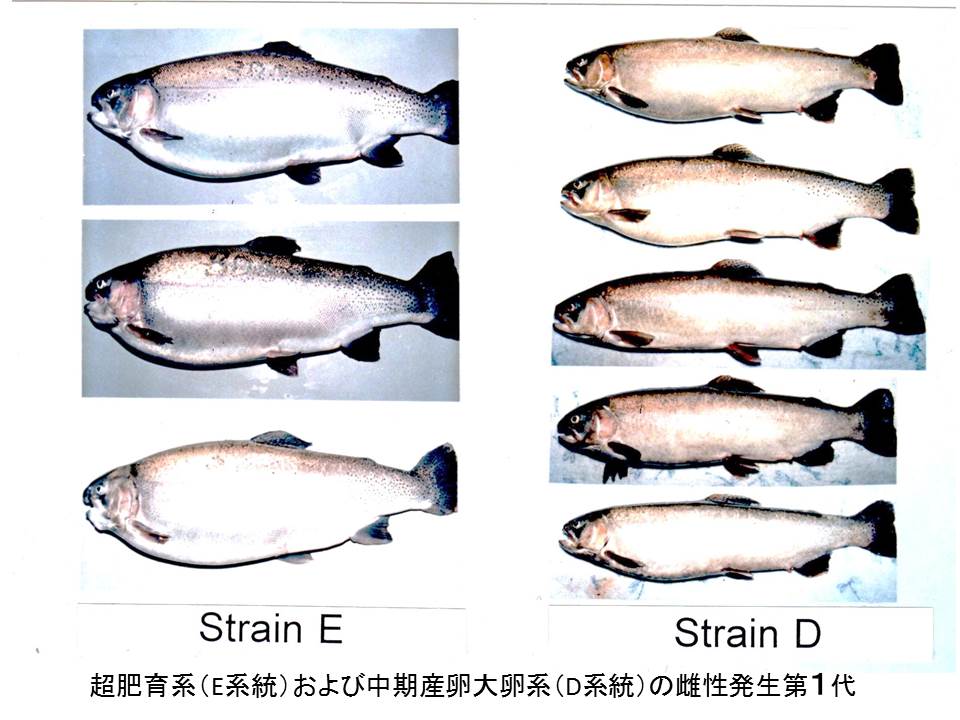
雌性発生二倍体作出法は、精子の遺伝的関与を除外できることから、雌親の特性のみを次世代に継承させ、有用な形質をもった系統を短期間(わずか2世代)で作出することができます。育種効果の起爆剤。発生初期の第2減数分裂や第一卵割の細胞分裂の阻止が、こんな効果をもたらします。また、第一卵割の阻止で作出した雌性発生二倍体の次世代はクローン(遺伝子構成が全く同じ個体群)となります。これらの方法を使えば、様々な特性をもった品種系統が短期間にたくさん作出できます。実用的な養殖品種として利用するためには、これらの雌性発生系統同士を交配したり、その中からさらに選抜したりして、丈夫でよりよい特性をもった系統をつくることが重要です。
優良新品種作出の可能性 −複二倍体ー
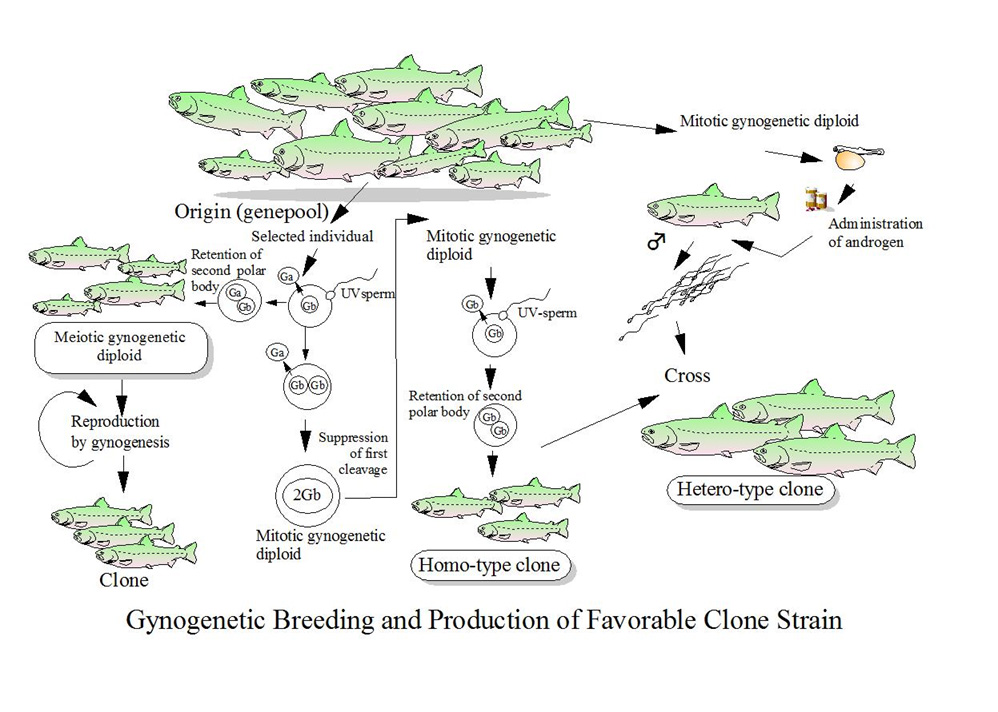 三倍体にしても、雌性発生二倍体にしても、やっていることは、希釈精液を紫外線にさらしたり、受精卵を氷水やぬるま湯に漬けたり、圧力をかけたりするだけです。しかし、それで起こる細胞分裂の阻止が、さまざまな倍数体を作り上げ、生物学的特性を大きく変化させます。これらを品種として利用することで、水産養殖業の発展に大きく貢献できると思います。
三倍体にしても、雌性発生二倍体にしても、やっていることは、希釈精液を紫外線にさらしたり、受精卵を氷水やぬるま湯に漬けたり、圧力をかけたりするだけです。しかし、それで起こる細胞分裂の阻止が、さまざまな倍数体を作り上げ、生物学的特性を大きく変化させます。これらを品種として利用することで、水産養殖業の発展に大きく貢献できると思います。
また、異種間での交雑では、ほとんどの場合不稔性を示し、系統化が出来ませんが、交雑卵の卵割阻止で作出が理論的に可能な複二倍体は、すべての染色体が2本ずつあることから、おそらく卵形成や精子形成がスムーズに行えるものができるでしょう。すなわち、雑種を系統化することも可能になるはずです。染色体操作には、まだまだ未知の領域がたくさんあります。
小林 徹 toru kobayashi
水産生物学研究室
Laboratory forAaquatic Biology
〒631-8505
奈良市中町3327-204